歩行のメカニズムを神経学的視点から解説
歩行のメカニズムについて以下の記事を書きました。
歩行分析のスキルアップをお考えの方はこちらの記事で分析に必要な知識を確認してみましょう。
運動学的視点から歩行のメカニズムをまとめた記事は下にあります。
この記事では歩行のメカニズムについて神経学的視点からまとめています。歩行を司る神経機構を理解することでリハビリの進め方が明確になりますので、最後までお読みください。
歩行の基本的神経基盤は皮質下構造による
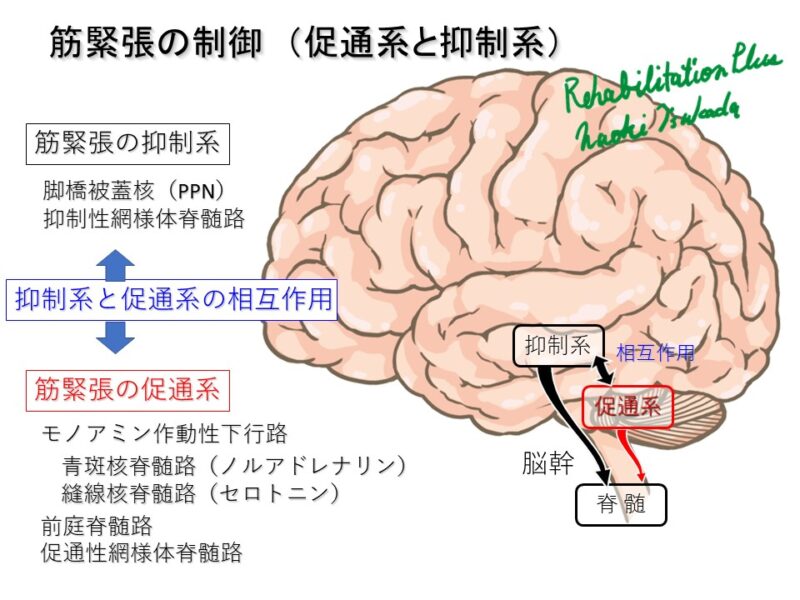
脳幹には筋のトーヌスを調整する脚橋被蓋核(抑制系)や網様体脊髄路(抑制系・促通系)、前庭脊髄路(促通系)などがあり、トーヌスを調整している。
これは除脳ネコの電気刺激により1990年代により証明されている。(森茂美, 1997, 運動の制御(Ⅳ)ブレインサイエンス)
歩行の神経機構
歩行の神経機構は主に上図の6つのエリアが関与しています。
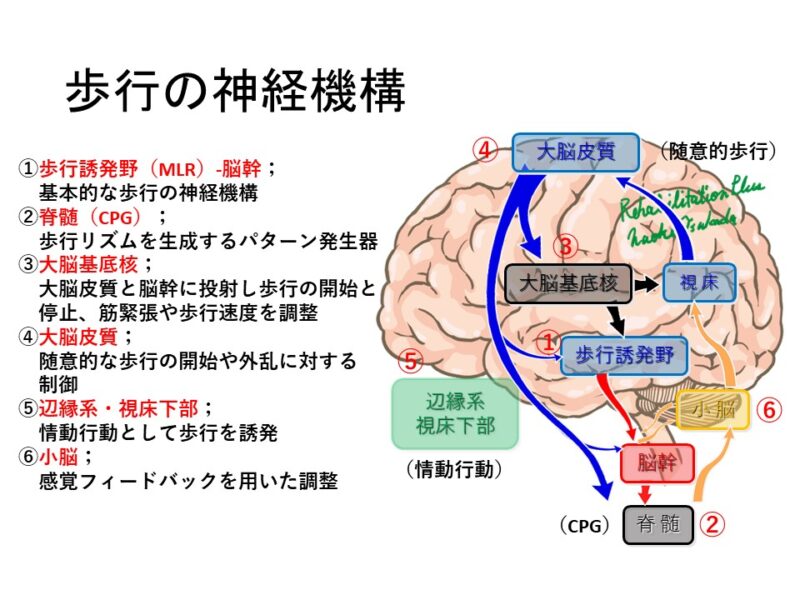
辺縁系で情動的に動き出す必要に迫られた場合⑤や、大脳皮質で目的を意図し④、基底核③で運動パターンを抽出すると、中脳の歩行誘発野①からの刺激は脊髄に下行しCPG②を発火させます。その結果は感覚のフィードバックとして小脳⑥へ入力され運動の修正が起こります。
これが歩行の開始から修正の一連の流れになります。
つまり、歩行の開始には意図がありますが、開始された歩行は皮質下の神経ユニットの働きにより継続されることになります。
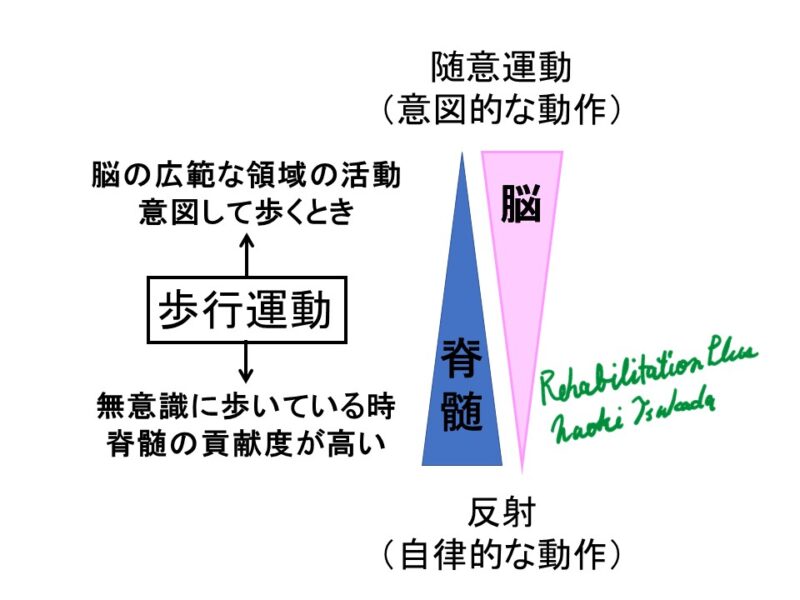
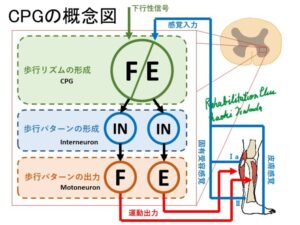
CPG(Central Pattern Generator)とは
歩行に必要な基本的運動パターンの生成は脳幹と脊髄のユニットによって構成されています。
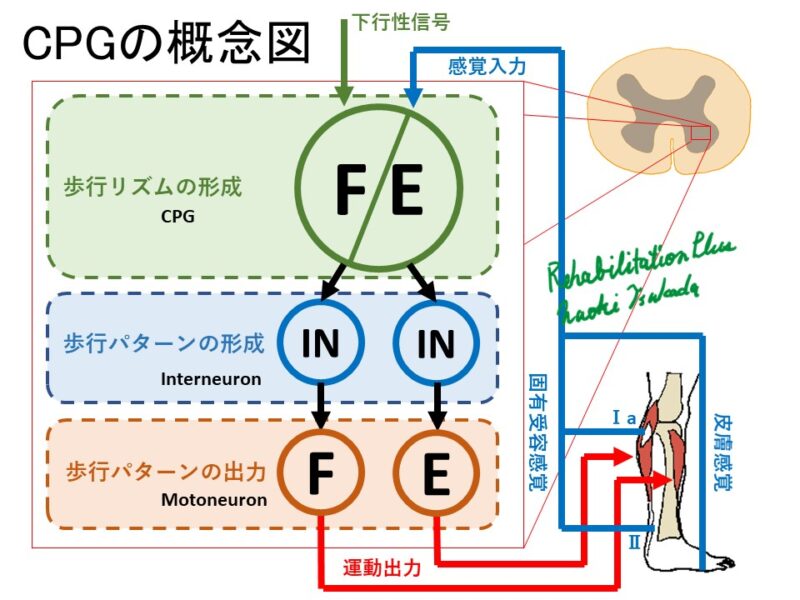
下行性シグナルを受け取った灰白質内の神経ユニットCPGは一側下肢の屈曲と伸展のリズムを生成します。
次いで伸展のHalf Centerが伸展のパターンを形成し、その後モーターニューロンプールにある運動ニューロンへパターンを出力しています。
屈曲のパターンを生成している場合は伸展のHalf Centerは抑制性介在ニューロンにより活動が抑えられています。
この機構により一側下肢の屈曲と伸展が作られています。
また対側下肢との連携は、灰白質Ⅷ層の介在ニューロンの多くが対側Ⅷ層に終始しておりこれにより連携していると考えられています。
リハビリでどう考えるか?
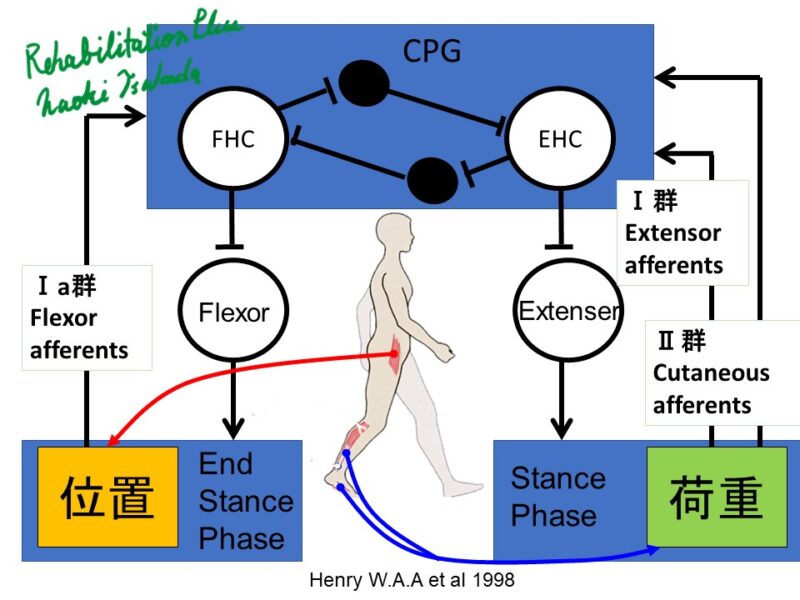
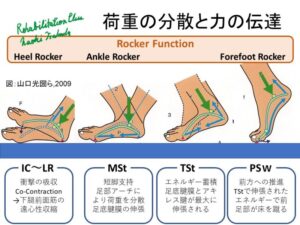
荷重情報を検出する主要な受容器には、アキレス腱、大腿四頭筋、足底筋のゴルジ腱器官 (Group Ⅰb)と足底の皮膚受容器(Group Ⅱ)があげられています。
(上林清孝, 2009, 歩行時の体性感覚の影響 皮質脊髄路興奮性に与える影響)
カンデルによると股関節屈筋が十分伸張されるまで、下肢の伸筋は興奮を続けます。(カンデル脳神経科学)
つまり、StanceからSwingへの切り替えは股関節屈筋が十分に伸張されることと荷重が踵から前足部に移ることが重要だと考えられます。
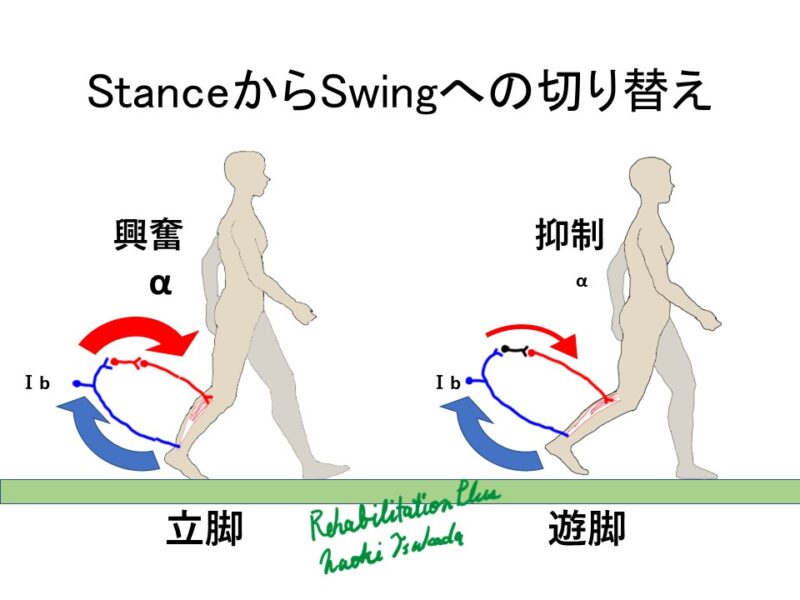
立脚相後期に股関節が伸展される際の筋紡錘からの求心性入力は、Swingへの位相転換を担う股関節屈筋群の活動を喚起します。(Grillner S and Rossignol S 1978)
エックマンによると(上図)足底が接地している間、同名筋に対して興奮性に働きます。これは、ゴルジ腱器官からのⅠb求心性情報によるとされています。
片麻痺患者はどうなっているか?
片麻痺の方を想像すると、次の二通りが多く見られます。
膝折れ:
この歩容を示す方は、立脚中の下肢の伸展が不十分ということになります。立脚中の下肢からのフィードバックが下肢の伸筋の興奮に及ぼす影響が乏しいことが一因と考えられます。
ぶん回し歩行:
この歩容を示す方は下肢の伸展から屈曲への切替ができていません。つまり、股関節が伸展し下肢が後ろに運ばれることがすくないと考えられます。
まとめ
歩行を作り出すの神経学的側面をまとめました。
リハビリを行う上でその他の考慮すべき事項については以下の記事にまとめています。
歩行のリハビリ全般のまとめ記事になります。